A mathematical model for period-memorizing behavior in Physarum plasmodium
Abstract
A mathematical model to describe period-memorizing behavior in Physarum plasmodium are reported. In constructing the model, we first examine the basic characteristics required for the class of models, then create a minimal linear model to fulfill these requirements. We also propose two modifications of the minimal model, nonlinearization and noise addition, which improve the reproducibility of experimental evidences. Differences in the mechanisms and in the reproducibility of experiments between our models and the previous models are discussed.
keywords:
amoeba locomotion, anticipation, integrate-and-fire model1 Introduction
The plasmodium of Physarum polycephalum is a huge unicellular organism with amoeboid movement. It is differentiated into an advancing front zone and a rear region that is composed of a network of protoplasmic veins. The concentrations of many chemical components in its cytoplasm, such as Ca2+(Ridgway et al.,, 1976; Yoshimoto et al., 1981a, ), ATP(Yoshimoto et al., 1981b, ), H+(Nakamura et al.,, 1982), cAMP(Ueda et al.,, 1986), and NADH(Mori et al.,, 1987), display mutually entrained oscillations in a period of 1-2 minutes. These oscillations also involve the contraction-relaxation cycle of cytoplasmic actomyosin(Wohlfahrt-Bottermann,, 1979), which generates rhythmic shrinkage of protoplasmic veins and the active transport of cytoplasm via shuttle streaming in the vein network. The streaming of cytoplasm pushes the front zone to spread out, and the solation of cytoplasmic gel in the zone occurs at the same time; thus the plasmodium migrates. This shuttle transport of cytoplasm also coordinates the phase relations among local oscillatory dynamics and lets a huge plasmodium behave as a unified individual organism(Yoshimoto et al.,, 1978).
Because of its particular behaviors, the plasmodium of P. polycephalum has been intensively examined as a model organism for the study of the cooperative phenomena in biological systems. Many theoretical approaches have also been reported, in which the plasmodium is modeled as spatially extended oscillatory medium (Teplov et al.,, 1991; Yamada et al.,, 1999) or coupled oscillator system(Akahane et al.,, 1999; Tero et al.,, 2005; Odell,, 1982; Smith,, 1994). A combination of the results of experimental and theoretical studies have revealed a lot of interesting features about plasmodium’s behaviors, such as entrainment to environmental stimulation(Miyake et al.,, 1992), mutual entrainment(Yoshimoto et al.,, 1978), symmetry-induced patterns(Takamatsu et al.,, 2001; Takamatsu,, 2006), maze-solving(Nakagaki et al.,, 2000), and computing(Aono et al.,, 2008).
Recently, another interesting behavior – memorization of the period of periodic stimulation – has been reported by Saigusa et al., (2008). In that study, the Physarum plasmodia were exposed to dry and cold condition (dry stimulation) three times periodically with periods - minutes. The stimulation reduced the locomotion speeds of the Physarum plasmodia. After the last stimulations, spontaneous slowdowns of locomotion speeds were observed. The time intervals between the last stimulations and spontaneous slowdowns coincided with the stimulation periods. This phenomenon was referred to as a spontaneous in-phase slowdown (SPS). The number of SPS occurrences variated from none to three, depending on the organism. Additionally at min after SPS stopped, one-time stimulation induced the spontaneous slowdowns after an equal period of time from the additional stimulation, which is referred to as SPS after one disappearance (SPSD). SPSD was not observed at min or min. These experiments clearly indicate that the Physarum plasmodium memorizes the period of the periodic stimulation. Furthermore, since the dry cold conditions used in the study are thought to be unfavorable for Physarum plasmodium, the spontaneous slowdown can be interpreted as anticipated behaviors against unfavorable conditions (Saigusa et al.,, 2008).
In this paper, we present mathematical models to explain this period-memorizing behavior. Since there is little information about the controlling mechanisms of the Physarum plasmodium’s locomotion, we assume a general dynamical system, and examine the basic characteristics required to control the plasmodium’s locomotion speed. We then present a simple model that fulfills these requirements and qualitatively reproduces experimental evidences. We insist the model is minimal in the sense that it has the minimum degrees of freedom and that the functions that it uses (including the differential equations to drive the system) are all linear functions. We also propose some modifications of the minimal model to improve the reproducibility of experimental evidence. Another mathematical model has been presented in a paper reporting these experiments (Saigusa et al.,, 2008), though that model’s mechanism differs from ours. We outline the previous model and compare it with our model in the Discussion section.
2 Basic requirements for models
In this section, we argue the class of models for a biochemical system which controls the reduction of the locomotion speed of the Physarum plasmodium. The main characteristics of the system are that the stimulation makes the system reduce the locomotion speed, and the system also spontaneously reduces the speed. Given that the speed reduction is a transient response, the systems behaviors are analogous to those of a neuron with steady and pulsing inputs. Thus, our model refers to simple neuron models: integrate-and-fire models (Burkitt,, 2006). We consider differential equations
| (1) |
to describe the autonomous dynamics of the biochemical system in P. polycephalum, and suppose that this system reduces the locomotion speed when the variables satisfy a condition
| (2) |
In other words, determines the threshold plane, and if the system gets across the plane, locomotion speed is reduced. If the stimulation is imposed, the orbit of the system in the sub-threshold region is forced to shift into the over-threshold region. On the other hand, if the orbit spontaneously crosses over the the threshold plane, SPS occurs.
In the following, we examine the experimental evidences, list four basic requirements for models, and give appropriate assumptions to meet these requirements.
Equilibrium states. Before the periodic stimulation was applied or long after stimulation was ceased, the plasmodium showed steady locomotion. Thus, the system should have equilibrium states in the sub-threshold region (), so that it does not reduce speed spontaneously in the equilibrium states.
Locomotion recovery. Physarum plasmodium recovered its locomotion speed after stimulation, and became ready to respond to the next stimulation. This implies that the condition eq. (2) does not hold for long after the slowdown begins. In order to bring the system’s state back to the sub-threshold region, here we assume that some of the variables are reset after the system induces the slowdown. Let be the variables to be reset and the variables not to be reset after the slowdown.
Transitive spontaneous action. SPS did not persist in the experiments. Thus, there should be a difference in the system’s actions between a stimulation-induced slowdown and spontaneous slowdown (SPS). We assume that the resetting manners differ. In the SPS case, the reset may start before the orbit invade deeply into the over-threshold region. However, the stimulation can be vigorous and may shift variables widely so that the function (2) takes a large positive value. This value may affect the resetting manner. In particular, we assume that after the stimulation, resettable variables are shifted to the complete reset state
| (3) |
On the other hand, the reset after SPS is incomplete: resettable variables are shifted to an internally dividing state between the state just before the reset and the complete reset state
| (4) |
Memory. The plasmodium remembered the stimulation period for a long time after stimulation stopped. In order for a dynamical system to memorize such an analog value, it must have a slow manifold in the phase space: the relaxation along with the manifold is quite slow so that the system can preserve the information about its own history for a long time (Fox, et al.,, 2002). As a simple and extreme case, we introduce the neutral manifold (a line of equilibrium states) in the phase space of our model, where the system never relaxes in that direction, and thus, the stored memory is permanently preserved. In other words, we introduce a conserved quantity whose value never changes in the autonomous dynamics. Along with those dynamics, the system’s orbit is constrained on a dimensional invariant plane transverse to the neutral manifold, and the resetting action will shift the orbit to another plane along with the neutral manifold. The period of stimulations is thought to control which plane is selected.
Summarizing the class of models, we can say that these models have a conservative quantity; thus the autonomous dynamics of the models are restricted to the dimensional invariant plane. In this plane there is an equilibrium state, a dimensional threshold plane, and a complete reset state. Changing the value of the conservative quantity changes the positional relation among them in the plane. The relation determines whether or not autonomous dynamics from the complete reset state to the equilibrium state cross the threshold plane, and if they do, it also determines the period of time from the complete reset state to the threshold plane.
3 Models and their behaviors
Based on the above discussions, here we present a minimal model for the slowdown induction system. The model has two resettable variables , , and one un-resettable variables , driven by linear differential equations,
| (8) |
where and indicate the relative time scales of , and respectively. These equations have a conserved quantity
| (9) |
Thus there are neutral manifold in the phase space, and the equilibrium states form a line
| (10) |
We set the threshold function as
| (11) |
where determines the distance between the equilibrium state and the threshold plane. We choose the complete reset state for resettable variables
| (12) |
Thus, reset after stimulation becomes
| (15) |
and reset after SPS becomes
| (18) |
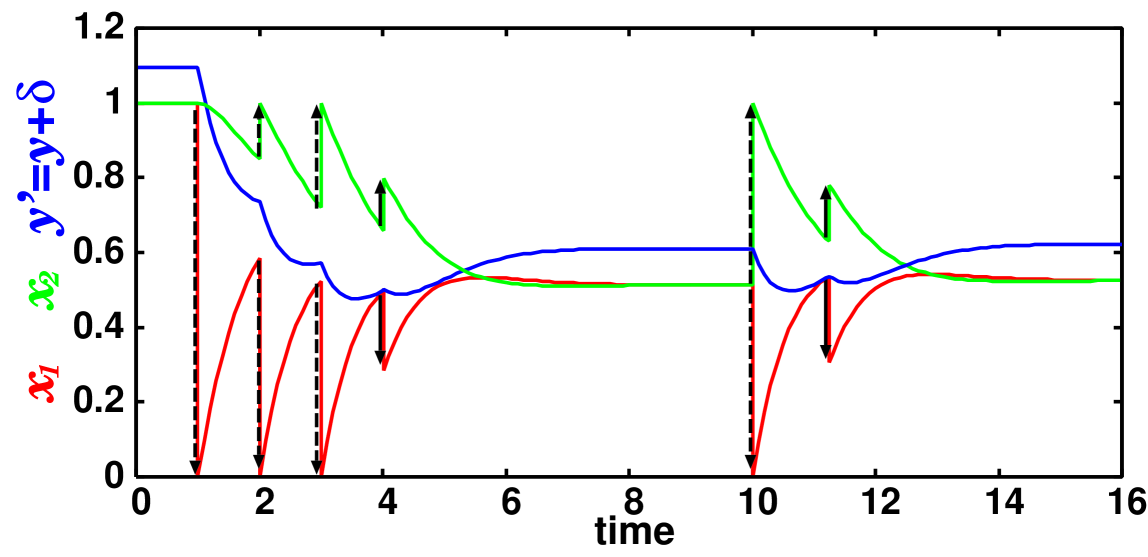
We show the behavior of our model in fig. 1. Initially the system stays at the equilibrium point . We give the stimulation three times. After that, at , the system reaches the point where eq. (11) changes the sign from negative to positive, SPS occurs, and the orbit relaxes to a new equilibrium point . After additional stimulation is imposed (), SPSD occurs at . and the orbit relaxes to an equilibrium point that is closer to the previous one ().
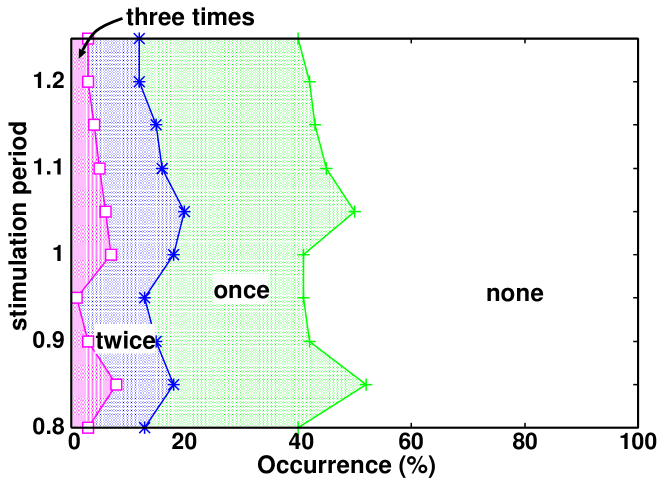
We repeated the simulations with various stimulation periods. The response periods of SPS and SPSD are shown in fig. 2. The parameters used in these simulations are tuned so that these response periods have similar values to the stimulation periods and show positive correlations as the stimulation period changes.
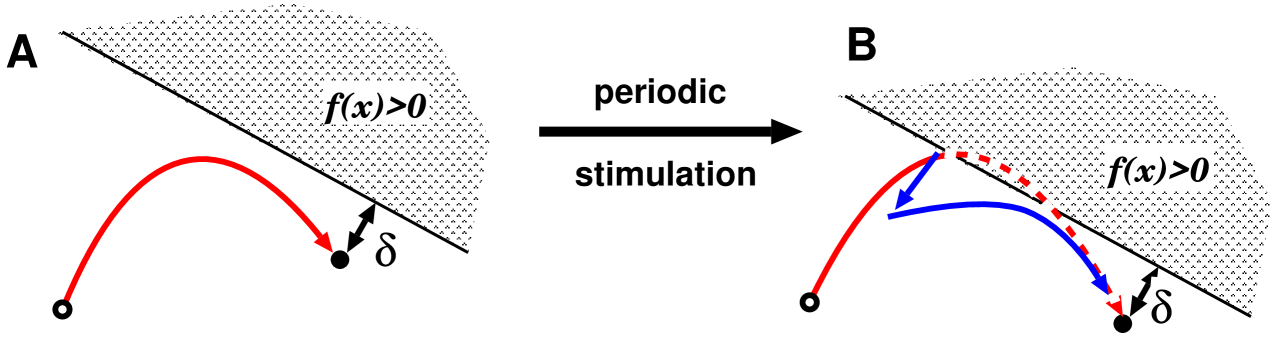
Here we sketch the spontaneous dynamics of the system, and verify that the above model with three variables is a minimal model. Because the system has one conserved quantity, the spontaneous dynamics are restricted in a two dimensional plane, which is described in fig. 3. The plane is divided by threshold line eq. (11) and has one equilibrium point and one complete reset state which lies in the sub-threshold region. Initially the arrangement of the plane may look like fig. 3-A. The relaxation orbit from the complete reset state directly relaxes to the equilibrium point. Periodic stimulation may change the positional relationship among the equilibrium point, the complete reset state, and the sub-threshold line, similar to fig. 3-B. That occurs due to the continuous change of the unresettable variable during the periodic stimulation. In the plane, the orbit from the complete reset state violates the threshold line in the course of relaxation to the equilibrium point, and thus SPS occurs. After the induction, the orbit shifts a bit toward the the complete reset state and relaxes to the equilibrium point. We emphasize that after periodic stimulation, the orbit from the complete reset state overreaches and crosses the threshold line, but the orbit after spontaneous induction directly relaxes to the equilibrium point. This happens because the plane is two-dimensional. If it is one-dimensional, there are only two cases: if the equilibrium point is in the sub-threshold region, the orbit relaxing to it never crosses the threshold, and if the equilibrium point is in the over-threshold region (this case is eliminated in the above assumptions), the orbit never reaches the equilibrium point. This ensures that the plane should be at least two-dimensional. Thus, with the neutral manifold for memory, a minimal model must have three variables.
To make the model minimal and easily understandable, we chose linear functions for differential equation eq. (8), the threshold function eq. (11) and resetting manner eq. (18). However, these functions can be nonlinear without changing the basic mechanisms. Here we give an example of the nonlinear modification of the model, by letting parameters , depend on variables
| (21) |
and changing the threshold function from eq. (11) to
| (22) |
where are the nonlinearity parameters. The modification improves the reproducibility of the periods, as displayed in fig. 2.
In experiments, the number of times of SPS occurrences varied among; whereas one organism showed SPS three times, another organism did not at all. These variations must be due to fluctuation in the cell state dynamics. Adding Langevin noise to the differential equations
| (26) |
where , our model also shows such varieties of responses, as shown in fig. 4.
4 discussion
In this paper, we presented a mathematical model for the period-memorizing behavior of the plasmodium of P. polycephalum. We described and examined the conditions for satisfying the model, and introduced a minimal linear model that qualitatively reproduces the experimental evidence. To improve the model, we also suggests the two methods (nonlinearization and noise addition) that are better for quantitative reproducibility.
Since almost nothing has been revealed about the chemical components for the locomotion speed reduction system of the Physarum plasmodium, we did not present a detailed model in which variables correspond to actual chemical components. Instead, we made a minimal linear model whose mechanism is easily comprehensible. In our model, for example, the system’s resetting action after it reduces the locomotion speed is modeled in a conditional branching manner, instead of in an autonomous dynamical system. In a detailed model, this process may be represented by neuron-like firing action. The abstraction revel of our model corresponds to those of integrate-and-fire models for a neuron (Burkitt,, 2006), as mentioned above. The transcriptome of P. polycephalum is now being sequenced (Glöchner, et al.,, 2008). This information may lead us to develop a more detailed model. In particular, two important actions for locomotion, actomyosin contraction and sol-gel transition of the cytoplasm, are thought to be controlled by the calcium ion concentration. Thus, transcripts homologous to what are involved in calcium ion concentration regulations in other organisms may give useful insight into the detailed mechanism.
Previously, another model for the plasmodium’s period-memorizing behavior was presented by Saigusa et al., (2008). Here we outline that model’s architecture and behaviors. That previous model consists of decoupled multiple oscillators. The oscillators form sub-groups, each of which contains a certain number of oscillators with the same frequency (). There are multiple sub-groups with different frequencies. The phase relations among oscillators are random at the initial condition. The periodic stimulation first synchronizes oscillators in phase within sub-groups that have similar periods to those in the stimulation, and then it synchronizes these sub-groups in phase. In that study, this super cluster (a cluster of clustered sub-groups) is assumed to induce SPS in their study. After stimulation is ceased, the super cluster gradually desynchronizes and SPSs are observed as only transient phenomena. However, the clusters within the sub-groups do not desynchronize, because they have exactly the same periods. An additional one-time stimulation again induces the formation of a super cluster, and thus induce SPSD. In the following, we remark on the two advantages of our model in reproduction of experimental evidence, and discuss two other prominent differences in the mechanism between the previous model and our model.
With the previous model, the occurrence of SPSD depends on at which phase the one-time stimulation is given; stimulation at the in-phase with previous periodic stimulation induces SPSD much more strongly than the stimulation at the anti-phase. However, this is not consistent with the experimental results. In the experiments, the occurrence of SPSD did not depend on a small shift in the timing of one-time stimulation 111the occurrence of SPSD seems to decrease monotonically when the delay in timing increases. This may be explained by the noise effect in both models. , which is reproduced in our model. Besides, SPSD was observed only at min in the experiments, and our model also gives SPSD in a certain window of the stimulation period ( with the linear model and with the nonlinear model). However, the occurrence of SPSD in the previous model does not depend on the stimulation period. Thus another mechanism will be required for the previous model to reproduce the occurrence of SPSD for a certain window of the stimulation period.
There are large differences in required degrees of freedom, between our model and the previous model. As described above, a response period in the previous model is represented by intrinsic collective modes of the dynamical system. This means that a number of variables (oscillators) are needed to respond to each stimulation period. Thus the model needs a lot of degrees of freedom. Actually, oscillators are used in the numerical simulation in the paper (Saigusa et al.,, 2008): it contains 440 sub-groups with different frequencies and each sub-group consists of 1000 oscillators. In our model, on the other hand, a response period is not generated by the intrinsic mode but is represented by the length in phase space between the reset state and the threshold plane, which is continuously adjustable. Thus our model shows qualitatively the same behaviors with only three degrees of freedom.
Because the previous model uses intrinsic modes, the stimulation period and response period show fairly good correspondence. No tuning of parameters is necessary for the correspondence. On the other hand, in our model, parameter tuning is necessary even for reproducing the qualitative behaviors. It should be noted that having a conserved quantity in a general dynamical system itself demands the tuning of parameters, since the system is structurally unstable (Guckenheimer, et al.,, 1983). However, if this period-memorizing qualification has an adaptive role as means for anticipating unfavorable conditions, it might be not so hard for biological organisms to tune these parameters in evolution. For example, the period and the response to the temperature changes of circadian oscillation systems are thought to be tuned evolutionarily in many organisms (Rosato et al.,, 1997).
5 Acknowledgments
The author is grateful to T. Nakagaki, A. Awazu, and S. Ishihara for providing the author with motivation to conduct this study.
References
- Akahane et al., (1999) Akahane, T., Miyake, Y., Yamaguchi, Y., 1999. Flexibility and consistency in adaptation of Physarum – Chemo-mechanical coupling model of pattern formation in taxis–. IEEE SMC ’99 Conference Proceedings, pp401-406.
- Aono et al., (2008) Aono, M., Hara, M., 2008. Spontaneous deadlock breaking on amoeba-based neurocomputer. BioSystems, 91, pp 83-93.
- Burkitt, (2006) Burkitt, A.N., 2006. A review of the integrate-and-fire neuron model: I. Homogeneous synaptic input. Biological Cybernetics, 95, pp 1-19.
- Fox, et al., (2002) Fox, J.J., Bodenschatz, E., GIlmour Jr., R.F., 2002. Period-doubling instability and memory in chardiac tissue. Physical Reciew Letters, 89, 138101.
- Glöchner, et al., (2008) Glöckner, G., Golderer, G., Werner-Felmayer, G., Meyer, S., Marwan, W., 2008. A first glimpse at the transcriptome of Physarum polycephalum. BMC Genomics, 9:6.
- Guckenheimer, et al., (1983) Guckenheimer, J. Holmes, P., Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields. Springer, New York (1983).
- Miyake et al., (1992) Miyake, Y., Yano, M., Tanaka, H., Shimizu, H., 1992. Entrainment to external Ca2+ oscillation in Ionophore-Treated Physarum plasmodium. Cell Structure and Function, 17, pp 371-375.
- Mori et al., (1987) Mori, Y., Ueda, T., Kobatake, Y., 1987. NAD(P)H oscillation in relation to the rhythmic contraction in the Physarum plasmodium. Protoplasma, 139, pp 141-144.
- Nakagaki et al., (2000) Nakagaki, T., Yamada, H., Toth A., ,2000. Maze-solving by an amoeboid organism. Nature, 407, pp 470.
- Nakamura et al., (1982) Nakamura, S., Yoshimoto, Y., Kamiya, N., 1982. Oscillation in surface pH of the Physarum plasmodium. Proc. Japan Acad 58 Ser B, pp 270-273.
- Odell, (1982) Odell, G.M., 1982. A mathematically modelled cytogel cortex exhibits periodic Ca2+-modulated contraction cycles seen in Physarum shuttle streaming. Journal of Embryology and Experimental Morphology, 83 Supplement, pp 261-287.
- Ridgway et al., (1976) Ridgway, E.B., Durham, A.C., 1976. Oscillation of calcium ion concentrations in Physarum polycephalum. The Journal of Cell Biology, 69, pp 223-226.
- Rosato et al., (1997) Rosato, E., Piccin, A., Kyriacou, C.P., 1997. Circadian rhythms: from behaviour to molecules. Bioessays, 19, pp 1075-1082.
- Saigusa et al., (2008) Saigusa, T., Tero, A., Nakagaki, T., Kuramoto, K., 2008. Amoebae anticipate periodic events. Physical Review Letters, 100, 018101.
- Smith, (1994) Smith, D.A., 1994. A local-oscillator theory of shuttle streaming in Physarum polycephalum. I: Kamiya’s double-chamber experiment. Protoplasma, 177, pp 163-170.
- Takamatsu et al., (2001) Takamatsu, A., Tanaka, R., Yamada, H., Nakagaki, T., Fujii, T., Endo, I., 2001. Spatio-temporal symmetry in rings of coupled biological oscillators if Physarum plasmodium. Physical Review Letters, 87, 078102.
- Takamatsu, (2006) Takamatsu, A., 2006. Spontaneous switching among multiple spatio-temporal patterns in three-oscillator systems constructed with oscillatory cells if true slime mold. Physica D, 223, pp 180-188.
- Teplov et al., (1991) Teplov, V.A., Romanovsky, Y.M., Latushkin, O.A., 1991. A continuum model of contraction waves and protoplasm streaming in strands of Physarum plasmodium. BioSystems, 24, pp 269-289.
- Tero et al., (2005) Tero, A., Kobayshi, R., Nakagaki, T., 2005. A coupled-oscillator model with a conservation law for the rhythmic amoeboid movements of plasmodial slime molds. Physica D. 205, pp 125-135.
- Ueda et al., (1986) Ueda, T., Matsumoto, K., Akitaya, T., Kobatake, Y., 1986. Spatial and temporal organization of intracellular adenine nucleotides and cyclic nucleotides in relation to rhythmic motility in Physarum plasmodium. Experimental Cell Research, 162, pp486-494.
- Wohlfahrt-Bottermann, (1979) Wohlfahrt-Bottermann, K.E., 1979. Oscillatory contraction activity in Physarum. Journal of Experimental Biology, 81, pp 15-32.
- Yamada et al., (1999) Yamada, H., Nakagaki, T., Ito, M., 1999. Pattern formation of a reaction-diffusion system with self-consitent flow in the amoeboid organism Physarum plasmodium. Physical Review E, 59, pp 1009-1014.
- Yoshimoto et al., (1978) Yoshimoto, Y., Kamiya, N., 1978. Studies on contraction rhythm of the plasmodial strand. III. Role of endoplasmic streaming in synchronization of local rhythm. Protoplasma, 95, pp 111-121.
- (24) Yoshimoto, Y., Matsumura, F., Kamiya, N., 1981. Simulataneous oscillations of Ca2+ efflux and tension generation in the permealized plasmodial strand of Physarum. Cell Motility, 1, pp 433-443.
- (25) Yoshimoto, Y., Sakai, T., Kamiya, N., 1981. ATP oscillation in Physarum plasmodium. Protoplasma, 109, pp 159-168.